Cấu tạo virus Cúm gia cầm
Xuất hiện từ năm 412 trước công nguyên nhưng mãi đến năm 1680 mới bắt đầu bùng phát thành đại dịch và từ đó đến nay, cúm gia cầm luôn là mối lo ngại của toàn thế giới. Bệnh diễn biến ngày càng phức tạp và gây ra hàng loạt thiệt hại không hề nhỏ trên động vật và cả con người bất chấp những tiến bộ vượt bậc trong lĩnh vực y học trong thế kỷ qua.
Trước những diễn biến phức tạp đó nhất là trong thời gian gần đây, chúng tôi quyết định cho ra đời loạt bài viết về cúm gia cầm bao gồm các thông tin chi tiết từ đặc điểm cấu tạo của virus, các chủng cúm đang lưu hành hiện nay, cách nhận diện cúm, cũng như cách thức phòng và kiểm soát cúm nhằm cung cấp cho người đọc những cái nhìn toàn diện nhất về bệnh cúm trên gia cầm.
Virus cúm có hình dạng và cấu tạo như thế nào?
Virus cúm gia cầm (hay còn gọi là virus cúm A, virus cúm gà) có tên khoa học là Avian Influenza (AI), thuộc họ Orthomyxoviridae trong hệ thống phân loại chung.
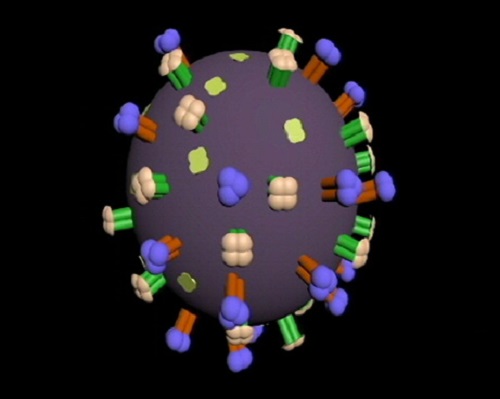
Virus cúm gia cầm
Virus cúm gia cầm có dạng hình cầu hay hình khối đa diện, đường kính từ 80-120nm. Đôi khi cũng có dạng hình sợi với chiều dài sợi có thể lên đến 2000nm và đường kính lõi khoảng 80-120nm. Virus có khối lượng phân tử là 250 triệu Da. Virus có cấu tạo đơn giản bao gồm vỏ (capsid), vỏ bọc ngoài (envelope) và lõi là ARN sợi đơn âm
Lớp vỏ virus
Lớp vỏ được bao bọc ngoài cùng bởi 1 lớp màng lipid kép có nguồn gốc từ màng tế bào nhiễm (thường là các tế bào trong đường hô hấp trên hoặc hồng cầu) đã được đặc hiệu hóa gắn các protein màng của virus cúm gia cầm.
Các protein màng (hay kháng nguyên bề mặt, hay gai glycoprotein) là các yếu tố kích thích sinh miễn dịch có ý nghĩa rất lớn đối với virus cúm gia cầm, chủ yếu bao gồm : HA, NA, protein kênh ion (M2), protein gian bào hay protein nền (M1).
Kháng nguyên bề mặt HA (Hemagglutinin)
HA (Hemagglutinin) là một glycoprotein thuộc protein màng typee I, có khả năng gây ngưng kết hồng cầu gà trong ống nghiệm .
Phân tử HA có dạng hình trụ, dài khoảng 130 ăngstron (Å), cấu tạo gồm 3 đơn phân (trimer), mỗi đơn phân (monomer) được tạo thành từ hai “dưới đơn vị HA1” và “ dưới đơn vị HA2” , liên kết với nhau bởi các cầu nối disulfide (-S-S-). Các đơn phân sau khi tổng hợp sẽ được glycosyl hóa (hay glycosylation là quá trình gắn kết các phân tử protein với nhau dưới tác động của enzym) như sau :
• “Dưới đơn vị HA1” sẽ gắn vào mặt ngoài vỏ capsid.
• “Dưới đơn vị HA2” (phần đầu tự do hình chỏm cầu) chứa đựng vị trí gắn với thụ thể thích hợp trên bề mặt màng tế bào đích.
Cấu tạo một phân tử HA
Trên bề mặt capsid của một virus có khoảng 400 phân tử HA với 2 chức năng chính là :
• Nhận diện và gắn virus cúm gia cầm với tế bào đích của vật chủ tương thích (ví dụ như các tế bào thuộc đường hô hấp trên và tế bào hồng cầu của động vật có xương sống).
• Sau đó khởi động quá trình xâm nhiễm của virus vào tế bào chủ đồng thời làm kết dính, ngưng kết hồng cầu hay dung hợp các tế bào bị nhiễm.
Có 18 nhóm HA (H1-H18), mỗi nhóm HA có khả năng kết hợp với một nhóm NA để tạo ra các chủng virus cúm gia cầm khác nhau, ví dụ như H5N6 (xem chi tiết cách phân chủng ở phần dưới).
Protein HA còn là kháng nguyên bề mặt quan trọng của virus cúm A và được coi là protein vừa quyết định tính kháng nguyên, vừa quyết định độc lực của virus. HA là đích của hệ thống miễn dịch, nó kích thích cơ thể sinh ra đáp ứng miễn dịch dịch thể đặc hiệu với từng typee HA , và tham gia vào phản ứng trung hòa virus, nhằm ngăn chặn sự xâm nhiễm của virus ở cơ thể nhiễm; từ đó là cơ sở điều chế các vaccine phòng cúm hiện nay.
Kháng nguyên bề mặt NA (Neurominidase)
Protein neurominidase còn gọi là sialidase, là một protein enzyme có bản chất là glycoprotein được gắn trên bề mặt capsid của virus cúm, mang tính kháng nguyên đặc trưng theo từng phân typee NA.
Có 9 phân typee (từ N1 đến N9) được phát hiện chủ yếu ở virus cúm gia cầm, hai phân typee N1 và N2 được tìm thấy ở virus cúm người liên quan đến các đại dịch cúm trong lịch sử (Webster, 1998). Có khoảng 100 phân tử NA xen giữa các phân tử HA trên bề mặt capsid virus.
Phân tử NA có dạng nút lồi hình nấm, đầu tự do (chứa vùng hoạt động) gồm 4 dưới đơn vị giống như hình cầu nằm trên cùng một mặt phẳng, và phần kị nước gắn vào vỏ capsid.
Cấu tạo 1 phân tử NA
Protein NA có vai trò là một enzyme cắt đứt liên kết giữa gốc sialic acid của màng tế bào nhiễm với phân tử cacbonhydrate của protein HA (sau khi HA kết dính vào bề mặt của tế bào nhiễm) giải phóng hạt virus cúm gia cầm ra khỏi màng tế bào nhiễm, đẩy nhanh sự lây nhiễm của virus trong cơ thể vật chủ, và ngăn cản sự tập hợp của các hạt virus mới trên màng tế bào.
Mặt khác, NA tham gia vào phân cắt liên kết này trong giai đoạn “hòa màng”, đẩy nhanh quá trình cởi áo “uncoating” giải phóng hệ gen của virus vào trong bào tương tế bào nhiễm, giúp cho quá trình nhân lên của virus diễn ra nhanh hơn.
Ngoài ra, NA còn phân cắt các liên kết glycoside, giải phóng neuraminic acid làm tan loãng màng nhầy bề mặt biểu mô đường hô hấp, tạo điều kiện cho virus nhanh chóng tiếp cận tế bào biểu mô và thoát khỏi các chất ức chế không đặc hiệu.
Cùng với vai trò của kháng nguyên HA, cả 3 khâu tác động trên của NA đều tham gia làm gia tăng độc lực gây bệnh của virus cúm A ở cơ thể vật chủ. Do đó, NA là đích tác động của các thuốc, hóa dược ức chế virus không đặc hiệu hiện nay, đặc biệt là Oseltamivir (biệt dược là Tamiflu) phong tỏa enzyme này, ngăn cản sự giải phóng hạt virus mới khỏi các tế bào đích, bảo vệ cơ thể (Theo Castrucci, Kawaoka ; Aoki và cộng sự).
Bên cạnh đó, NA còn là một kháng nguyên bề mặt của virus cúm gia cầm, tham gia kích thích hệ thống miễn dịch của cơ thể vật chủ, sinh ra kháng thể đặc hiệu với kháng nguyên NA của các chủng virus đương nhiễm có tác dụng phong tỏa protein NA (Theo Doherty và cộng sự, 2006).
Như vậy, kháng nguyên NA cùng với kháng nguyên HA của virus là các đích chủ yếu của cơ chế bảo hộ miễn dịch của cơ thể với virus cúm gia cầm, và là cơ sở điều chế các vaccine phòng cúm hiện nay cho người và gia cầm, nhằm ngăn chặn dịch cúm ở gia cầm và hạn chế lây truyền sang người.
Protein kênh ion (M2)
Protein M2 bao gồm 3 phân đoạn protein với tổng cộng là 97 amino acid.
M2 là một kênh proton, nghĩa là nó điều hòa sự vận chuyển các ion H+ đi qua màng virus khi điều kiện PH môi trường có sự thay đổi. Cụ thể, khi virus cúm gia cầm đi vào trong một tế bào vật chủ, PH môi trường giảm xuống >> kích thích kênh M2 mở ra và các proton (H+) từ nguyên sinh chất của tế bào vật chủ đi qua màng vào trong virus, điều này làm khởi phát quá trình phân ly của vỏ virus và cho phép các ARN của virus tự do phiên mã thành protein virus.
Protein nền (M1)
Protein M1 là một trong số các protein ma trận của virus cúm gia cầm. Tất cả các phân tử M1 xếp sát với nhau tạo thành một lớp áo khoác bên trong lớp màng lipid và là cầu nối giữa lớp màng với nhân ARN của virus. Đồng thời nó cũng giữ những chức năng quan trọng trong quá trình tái tạo, lắp ráp và nhân lên của virus cúm gia cầm như:
- Vận chuyển Ribonucleoprotein (RNP) từ ngoài vào trong và từ trong ra khỏi nhân virus cúm gia cầm. Cụ thể, khi virus xâm nhập vào tế bào vật chủ rồi nhân bản lên thành nhiều đoạn RNP, M1 sẽ liên kết với RNP mới đó và di chuyển lần lượt các RNP đó vào trong nhân virus mới (M1 sẽ tách khỏi RNP sau khi vận chuyển RNP đến đích). Trong giai đoạn cuối của quá trình nhiễm trùng, M1 ức chế hoạt động của ARNvirus bằng cách gắn vào RNP → tạo nên tín hiệu cho quá trình vận chuyển RNP từ trong nhân ra bề mặt tế bào. Cuối cùng, M1 kết hợp giữa RNP với glycoprotein của vỏ virus trên bề mặt bên trong của màng tế bào chất, sau đó thúc đẩy sự hình thành và nhân lên của virus cúm gia cầm.
- Lắp ráp các bộ phận của virus cúm gia cầm thành 1 con virus hoàn chỉnh (nó giống như 1 “chiếc xe vận chuyển”).
- Sự tương tác của M1 với các protein vỏ virus là rất cần thiết để chỉ đạo M1 đến màng tế bào phóng thích RNP (nhân) của virus cúm gia cầm.
Nhân ARN
Virus cúm gia cầm có nhân là ARN sợi đơn âm bao gồm 8 phân đoạn gen riêng biệt mang tên từ 1-8 theo thứ tự giảm dần của kích thước phân tử (hay được gọi theo tên protein mà chúng mã hóa tổng hợp), mã hóa cho 11 protein khác nhau là HA, NA, NP, M1, M2, PB1, PB1 – F2, PB2, PA, NS1 và NS2.
Mỗi phân đoạn RNA của virus cúm gia cầm có cấu trúc xoắn bậc 2 α đối xứng dài 50 - 100 nm, đường kính 9 - 10 nm, được bao bọc bởi nucleoprotein (NP) - bản chất là lipoprotein, tạo thành cấu trúc ribonucleoprotein (RNP). Mỗi RNP kết hợp với 3 protein enzyme polymerase (PA, PB1 và PB2) chịu trách nhiệm trong quá trình phiên mã và sao chép RNA của virus (Hình 5).
Các phân đoạn của hệ gen virus cúm A nối với nhau bằng các cầu nối peptide tạo nên vòm (loop) tại giới hạn cuối của mỗi phân đoạn, và tạo thành một sợi RNA duy nhất có tổng độ dài từ 10.000 - 15.000 bp (tuỳ theo từng chủng virus cúm gia cầm), chứa đựng khoảng 13,5 kilobase thông tin di truyền và có cấu trúc xoắn α bên trong vỏ virus.
Cấu tạo chi tiết của virus cúm và hệ gen
8 đoạn ARN đươc bố trí trong nhân virus
Sau đây là chi tiết đặc điểm của 8 phân đoạn gen của virus cúm gia cầm:
Tại sao virus cúm gia cầm lại nguy hiểm như vậy?
Thứ nhất, hệ gen của virus cúm gia cầm luôn luôn biến đổi rất linh hoạt tạo thành các chủng cúm mới có khả năng thích nghi cao hơn và khả năng gây bệnh nặng hơn chủng cũ.
Thứ hai, virus cúm gia cầm có hệ gen được cấu trúc từ 8 phân đoạn riêng biệt và không có gen mã hóa enzyme sửa chữa RNA, tạo điều kiện thuận lợi cho sự xuất hiện các đột biến điểm trong các phân đoạn gen/hệ gen qua quá trình sao chép nhân lên của virus, hoặc trao đổi các phân đoạn gen giữa các chủng virus cúm đồng nhiễm trên cùng một tế bào, rất có thể dẫn đến thay đổi đặc tính kháng nguyên tạo nên các chủng viruscúm gia cầm mới.
Thứ ba, virus cúm gia cầm độc lực cao có sự thích nghi rất tốt trên nhiều loại vật chủ khác nhau như gia cầm, động vật có vú, và con người. Điều đó làm cho việc kiểm soát mầm bệnh là vô cùng khó khăn.
Thứ tư, đa phần virus khu trú trong cơ thể chim hoang dã nhưng không có biểu hiện triệu chứng bệnh, điều đó đồng nghĩa với việc chúng ta gần như không thể kiểm soát được sự phát tán của mầm bệnh.
Cách đặt tên cho các chủng virus cúm gia cầm:
Virus cúm có 3 type A, B, và C khác nhau:
- Type A gây cúm ở các loại gia cầm: gà, gà tây, ngan, vịt, ngỗng, vẹt, cú, sẻ, bồ câu, lợn, ngựa, người ...
- Type B gây bệnh cúm ở người và ngựa.
- Type C chủ yếu gây bệnh cúm ở trẻ nhỏ không gây thành dịch.
Trong đó type A là type gây bệnh phổ biến và nguy hiểm nhất. Trong 15 nhóm thuộc type A thì H5 và H7 có nhiều chủng độc lực cao nhất.
Người ta phân loại virus cúm type A thành các subtype dựa trên các kháng nguyên bề mặt HA (1-18) và NA (1- 9).
Ví dụ như: chủng H7N6 tức là trên vỏ bọc của virus chỉ có gai H7 và gai N9. Từ đó, đáp ứng miễn dịch giữa các chủng virus cúm gia cầm trên cơ thể nhiễm bệnh là khác nhau.
Có 18 phân typee HA và 9 phân typee NA đã được phát hiện, sự tổ hợp giữa các phân typee này, về lí thuyết, có thể tạo ra rất nhiều các biến chủng khác nhau.
Khả năng gây bệnh của virus cúm gia cầm:
Virus cúm gia cầm có tính thích ứng lây nhiễm cao với biểu mô đường hô hấp, gây bệnh chủ yếu ở đường hô hấp, và cũng có thể tác động gây tổn thương nhiều cơ quan khác trong cơ thể của các động vật cảm nhiễm, do đó còn được gọi là virus hướng đa phủ tạng.
Khả năng gây bệnh của virus cúm A phụ thuộc vào độc lực và tính thích nghi vật chủ của từng chủng virus. Thông thường chúng không gây bệnh hoặc chỉ gây bệnh nhẹ giới hạn ở đường hô hấp của chim hoang dã và gia cầm nhiễm, nhưng một số chủng cường độc (H5, H7, và H1, H2, H3) có thể gây bệnh nặng ở hầu hết các cơ quan trong cơ thể, gây nên dịch cúm ở gia cầm và ở người, có lẽ do tính thích ứng thụ thể sialic của chúng.
Sau khi bị nhiễm virus cúm gia cầm, cơ thể vật chủ sinh ra đáp ứng miễn dịch chống lại virus bảo vệ cơ thể, nhưng đáp ứng miễn dịch này có thể không có tác dụng bảo vệ hoàn toàn cho những lần nhiễm sau, do virus cúm gia cầm luôn có sự biến đổi kháng nguyên của nó trong quá trình lưu hành ở tự nhiên, và không có đáp ứng miễn dịch chéo giữa các chủng virus cúm gia cầm (Webster, 1998). Do đó, khi xuất hiện những biến chủng virus cúm gia cầm có đặc tính kháng nguyên khác với các chủng virus trước đó, cơ thể nhiễm sẽ không hoặc ít có đáp ứng miễn dịch bảo hộ thích ứng với chủng virus cúm mới.
Đây là nguyên nhân làm cho gia cầm và con người thường bị mắc bệnh cúm nhiều lần trong năm, và các đợt dịch cúm xảy ra về sau thường nặng nề hơn và có thể gây nên đại dịch cúm mới.
Khả năng gây bệnh của biến chủng virus cúm gia cầm mới giảm hoặc biến mất, khi cơ thể có được đáp ứng miễn dịch đặc hiệu với biến chủng đó và chúng trở nên thích nghi lây nhiễm ở loài vật chủ mới. ví dụ: virus A/H1N1, A/H2N2, A/H3N2 là nguyên nhân của các đại dịch cúm trên người trước đây và đã thích nghi lây nhiễm ở người. Tuy nhiên, các chủng này vẫn thường gây ra các vụ dịch cúm tản phát hàng năm ở người, do khả năng biến đổi kháng nguyên của chúng. Đây cũng chính là nguồn virus trao đổi gen với các chủng virus cúm đang lưu hành ở gia cầm, để thích ứng lây nhiễm gây bệnh cho nhiều loài khác ngay cả trên người.
Độc lực gây bệnh của virus cúm gia cầm
Tính gây bệnh hay độc lực của virus cúm gia cầm được chia làm hai loại: Loại độc lực cao (HPAI - Highly pathogenic avian influenza), và loại độc lực thấp (LPAI - Low pathogenic avian influenza), cả hai loại đều cùng tồn tại trong tự nhiên.
- HPAI: là loại virus cúm A có khả năng gây tổn thương nhiều cơ quan nội tạng trong cơ thể nhiễm, trên gia cầm chúng thường gây chết 100% số gia cầm bị nhiễm trong vòng 48h sau nhiễm. Loại này rất nguy hiểm gây lo ngại cho cộng đồng. Virus loại HPAI phát triển tốt trên tế bào phôi gà và tế bào thận chó trong môi trường nuôi cấy không có trypsin.
- LPAI: là loại virus khi phát triển trong cơ thể nhiễm, có thể gây bệnh cúm nhẹ không có triệu chứng lâm sàng điển hình và không làm chết vật chủ. Đây là loại virus lây truyền rộng rãi và tạo nên các ổ bệnh trong tự nhiên của virus cúm A, loại này có thể trao đổi gen với các chủng virus có độc lực cao đồng nhiễm trên cùng một tế bào, và trở thành loại virus HPAI nguy hiểm.
Tính thích ứng đa vật chủ của virus cúm gia cầm
Vật chủ tự nhiên của tất cả các chủng virus cúm gia cầm là chim hoang dã (chủ yếu là vịt trời), đây là nguyên nhân lan truyền virus trong tự nhiên rất khó kiểm soát. Virus cúm có khả năng gia tăng loại vật chủ của chúng trong quá trình lây truyền ở tự nhiên.
Nhờ đặc tính luôn thay đổi kháng nguyên trong tự nhiên, virus cúm gia cầm có khả năng xâm nhiễm ở nhiều loài vật chủ trung gian khác nhau như gia cầm, một số loài động vật có vú (hải cẩu, cá voi, ngựa, lợn) và cả ở người, tạo nên tính thích ứng lan truyền “nội loài” như gà - gà, hay “ngoại loài” như gà - lợn; gà - lợn - người. Vịt (vịt trời) và một số loài thuỷ cầm khác (ngỗng) luôn luôn là vật chủ tàng trữ nguồn virus gây nhiễm.
Đặc điểm thích ứng vật chủ này là điều kiện thuận lợi cho virus cúm A trao đổi, tái tổ hợp các phân đoạn gen, đặc biệt là các phân đoạn gen kháng nguyên (gen “độc” HA và NA) giữa các chủng, tạo ra một chủng virus cúm mới có khả năng thích ứng xâm nhiễm ở loài vật chủ mới của chúng đặc biệt khi chúng vượt qua được “rào cản loài” dễ dàng thích ứng lây nhiễm gây bệnh từ gia cầm sang người và giữa người với người.
Trong lịch sử các đại dịch cúm ở người, lợn thường là vật chủ trung gian chuyển tiếp giúp cho virus cúm Gia cầm biến đổi để dễ dàng lây nhiễm sang người gây nên bệnh dịch. Ví dụ: cúm A/H3N2 là kết quả tái tổ hợp tự nhiên của virus cúm A/H2N2 của người và virus chứa gen H3 trong tự nhiên thông qua đồng nhiễm trên lợn, gây nên đại dịch cúm châu Á năm 1968.
Virus cúm A tương đối nhạy cảm với các tác nhân bất hoạt vật lí hay hóa học. Các hạt virus tồn tại thích hợp trong khoảng pH từ 6,5 đến 7,9. Ở pH quá acid hay quá kiềm, khả năng lây nhiễm của virus bị giảm mạnh.
Lớp vỏ ngoài của virus bản chất là lớp lipid kép, có nguồn gốc từ màng tế bào nhiễm, dễ bị phá hủy bởi các dung môi hòa tan lipid, chất tẩy rửa và các chất sát trùng: formaldehyde, phenol, β-propiolacton, sodium hypochloride, acid loãng và hydroxylamine. Virus bị bất hoạt dưới ánh sáng trực tiếp sau 40 giờ, tồn tại được 15 ngày ánh sáng thường, tia tử ngoại bất hoạt được virus nhưng không phá hủy được kháng nguyên của virus cúm gia cầm.
Tuy nhiên, virus cúm A dễ dàng bị tiêu diệt hoàn toàn ở 100oC và ở 60oC/30 phút, tồn tại ít nhất 3 tháng ở nhiệt độ thấp (trong phân gia cầm), và tới hàng năm ở nhiệt độ bảo quản (−70oC). Trong phủ tạng gia cầm (40oC), virus tồn tại 25 - 30 ngày, nhưng chỉ tồn tại 7 - 8 ngày ở nhiệt độ cơ thể người (37oC); trong nước, virus có thể sống tới 4 ngày ở nhiệt độ 30 độ C.
Như vậy, việc nắm rõ cấu trúc, cũng như những đặc tính cơ bản của virus cúm gia cầm sẽ giúp chúng ta phần nào hiểu rõ hơn mức độ nguy hiểm của bệnh đồng thời nó cũng là cơ sở để chúng ta đưa ra các phương án tốt nhất để khắc chế virus, nhằm giảm thiểu mức tối đa thiệt hại về kinh tế và đời sống con người do bệnh cúm gia cầm gây ra.